Общие положения
Замечание 1
Компоненты иммунной системы включают разнообразные клетки, ткани и органы, обеспечивающие иммунную защиту организма.
К иммунной системе относятся:
- центральные органы (тимус и костный мозг);
- периферические системы и органы (лимфатические узлы и лимфоидные скопления в различных органах, селезенка);
- пути циркуляции иммунокомпетентных клеток.
Помимо перечисленных органов в состав иммунной системы входят:
- миндалины носоглотки,
- Пейеровы бляшки кишечника,
- лимфоидные узелки слизистой оболочки дыхательной трубки, желудочно-кишечного тракта, урогенитальных путей,
- лимфоидные клетки Lamina propria,
- диффузная лимфоидная ткань,
- межэпителиальные лимфоциты.
В состав иммунной системы входят гуморальные факторы, растворимые молекулы, являющиеся продуктом В-лимфоцитов (антитела , иммуноглобулины) и цитокины – растворимые медиаторы межклеточных взаимодействий.
Органно-циркуляторный принцип организации иммунной системы
Лимфоидные клетки являются главным элементом иммунной системы.
При обеспечении иммунной функции организма лимфатическая система тесно взаимодействует с кровеносной системой, кожей и слизистыми оболочками, а также с другими органами.
Примерно каждая десятая клетка в организме человека является лимфоцитом.
Замечание 2
По анатомо-физиологическому принципу иммунная система устроена органно-циркуляторно, то есть лимфоциты постоянно циркулируют между нелимфоидными тканями и лимфоидными органами через лимфатические сосуды и кровь.
Передвижение лимфоцитов обеспечивают специфические взаимодействия молекул на мембранах лимфоцитов и эндотелиальных клеток сосудистой стенки. Данные молекулы:
- адгезины,
- интегрины,
- селектины,
- хоминг-рецепторы.
Благодаря этому каждый орган имеет специфический набор популяций лимфоцитов и клеток-партнеров.
Состав иммунной системы
Иммунная система состоит из разнообразных тканей и органов:
- кроветворный костный мозг;
- инкапсулированные органы (тимус, лимфатические узлы, селезенка);
- неинкапсулированная лимфоидная ткань (Пейеровы бляшки тонкого кишечника, лимфоидное глоточное кольцо Пирогова-Вальдейера, лимфоидная ткань слизистых оболочек бронхов и бронхиол, желудка и кишечника, органов мочеполовой системы и др.);
- периферическая кровь, выполняющая функцию транспортно-коммуникационного компонента иммунной системы.
В иммунной системе выделяют:
- Центральные органы. В кроветворном костном мозге и тимусе происходит дифференцировка моноцитов и лимфоцитов (миелопоэз, лимфопоэз).
- Периферические органы: лимфатические узлы, неинкапсулированная лимфоидная ткань, селезенка. В этих органах антигенраспознающие клетки взаимодействуют со зрелыми наивными лимфоцитами. В них происходит иммуногенез – додифференцировка лимфоцитов, в результате чего образуются клоны эффекторных лимфоцитов, которые способны распознать антиген и осуществить его деструкцию и периферических тканей организма, содержащих этот антиген.
Клетки иммунной системы
Иммунная система содержит клетки различного генеза:
- Клетки мезенхимного происхождения: все разновидности лимфоцитов, или собственно иммуноциты (Т-клетки, В-клетки, NK-клетки). Во время иммунного ответа эти клетки кооперируются с лейкоцитами (макрофагами/моноцитами, эозинофилами, нейтрофилами, базофилами, эндотелиоцитами сосудов, тучными клетками). Эритроциты транспортируют в селезенку и печень иммунные комплексы «антиген-антитело-комплемент» для фагоцитоза и разрушения.
- Эпителий. Некоторые лимфоидные органы содержат в своем составе клетки энтодермального и эктодермального происхождения.
Ярко-красного цвета, непрерывно циркулирующая по замкнутой системе кровеносных сосудов . В организме взрослого человека содержится приблизительно 5 литров крови. Часть крови (около 40 %) не циркулирует по кровеносным сосудам, а находится в «депо» (капиллярах, печени, селезенке, легких, коже). Это резерв, поступающий в кровяное русло в случае кровопотери, мышечной работы или недостатка кислорода. Кровь имеет слабощелочную реакцию.
Кровь
Клетки (46 %) – форменные элементы: эритроциты, лейкоциты, тромбоциты;
Плазма (54 %) – жидкое межклеточное вещество = вода + сухое вещество (8–10 %): органические вещества (78 %) – белки (фибриноген, альбумин, глобулины), углеводы, жиры; Неорганические вещества (0,9 %) – минеральные соли в виде ионов (К+, Na+, Ca2+)
Плазма – бледно-желтая жидкость, в состав которой входит вода (90 %) и растворенные, взвешенные в ней вещества (10 %); представляет собой кровь, очищенную от клеток крови (форменных элементов).
Кроме воды в состав плазмы входят разнообразные вещества, основу которых составляют белки: сывороточный альбумин, связывающий кальций, сывороточные глобулины, выполняющие функции переноса веществ и осуществления иммунных реакций; протромбин и фибриноген, участвующий в процессах метаболизма. Кроме того, в плазме содержится большое количество ионов, витамины, гормоны, растворимые продукты пищеварения и вещества, образовавшиеся в процессе метаболических реакций. Кроме того, из плазмы можно выделить сыворотку. Сыворотка почти тождественна плазме по составу, но в ней отсутствует фибриноген. Образуется сыворотка при свертывании крови вне организма после отделения от нее кровяного сгустка.
Форменными элементами крови являются:
Эритроциты
– мелкие безъядерные клетки двояковогнутой формы. Они имеют красный цвет из-за присутствия белка – гемоглобина, состоящего из двух частей: белковой – глобина и железосодержащей – гема. Эритроциты образуются в красном костном мозге и переносят кислород ко всем клеткам. Открыты эритроциты Левенгуком в 1673 году. Количество эритроцитов в крови взрослого человека составляет 4,5–5 млн. на 1 кубическом мм. В состав эритроцитов входит вода (60 %) и сухой остаток (40 %). Кроме переноса кислорода эритроциты регулируют количество различных ионов в плазме крови, участвуют в гликолизе, отбирают на себя токсины, и некоторые лекарственные вещества из плазмы крови, фиксируют некоторые вирусы.
Среднее содержание гемоглобина в 100 г. крови у здоровых женщин составляет 13,5 г., а у мужчин – 15 г. Если выделенную из организма кровь с предохраняющей от свертывания жидкостью поместить в стеклянный капилляр, то эритроциты начнут склеиваться и оседать на дно. Это принято называть скоростью оседания эритроцитов (СОЭ). В норме СОЭ составляет 4–11 мм./ч. СОЭ служит важным диагностическим фактором в медицине.

Лейкоциты
– бесцветные ядерные клетки крови человека. В покое имеют округлую форму, способны активно передвигаться, могут проникать сквозь стенки сосудов. Основная функция – защитная, с помощью ложноножек поглощают и уничтожают различные микроорганизмы. Лейкоциты также были открыты Левенгуком в 1673 году и классифицированы Р. Вирховым в 1946 году. Различные лейкоциты имеют в составе цитоплазмы гранулы, либо не имеют, но в отличие от эритроцитов, имеют ядро.
Гранулоциты. Образуются в красном костном мозге. Имеют разделенное на лопасти ядро. Способны к амебоидному движению. Подразделяются на: нейтрофилы, эозинофилы, базофилы.
Нейтрофилы . Или фагоциты. На их долю приходится около 70 % всех лейкоцитов. Они проходят пространства между клетками, образующими стенки сосудов, и направляются к тем участкам тела, где обнаруживается очаг внешней инфекции. Нейтрофилы являются активными поглотителями болезнетворных бактерий, которых переваривают внутри образующихся при этом лизосом.
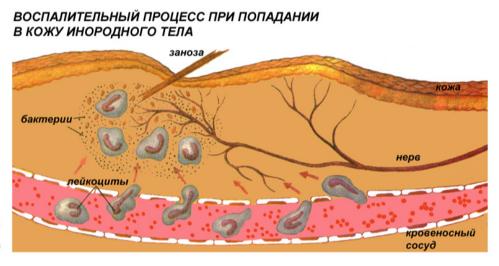
Тромбоциты – самые мелкие клетки крови. Их иногда называют кровяными пластинками, они безъядерные. Главная функция – участие в свертывании крови. Тромбоциты называют кровяными пластинками. По сути своей клетками не являются. Представляют собой обломки крупных, содержащихся в красном костном мозге клеток – мегакариоцитов. В 1 мм3 крови взрослого человека содержится 230–250 тыс. тромбоцитов.
Функции крови:
Транспортная – кровь переносит кислород, питательные вещества, удаляет углекислый газ, продукты обмена, распределяет тепло;
Защитная – лейкоциты, антитела защищают от инородных тел и веществ;
Регуляторная – по крови распространяются гормоны (вещества, регулирующие жизненно-важные процессы);
Терморегулирующая – кровь переносит тепло;
Механическая – придает органам упругость за счет прилива крови.
Иммунитет – способность организма защищать себя от болезнетворных микробов и , инородных тел и веществ.
Иммунитет бывает:
Естественный – Врожденный, Приобретенный
Искусственный – Активный (вакцинация), Пассивный (введение лечебной сыворотки)
Защита организма от инфекции осуществляется не только клетками – фагоцитами, но и особыми белковыми веществами – . Физиологическую сущность иммунитета определяют две группы лимфоцитов: Б– и Т–лимфоциты. Важным является укрепление естественного врожденного иммунитета. У человека выделяют два вида иммунитета: клеточный и гуморальный. Клеточный иммунитет связан с наличием в организме Т–лимфоцитов, которые способны связываться с антигенами чужеродных частиц и вызывать их разрушение.
Гуморальный иммуните
т связан с наличием В–лимфоцитов. Эти клетки выделяют химические вещества – антитела. Антитела, присоединяясь к антигенам ускоряют их захват фагоцитами, либо приводят к химическому разрушению или склеиванию и осаждению антигенов.
Естественный врожденный иммунитет
. В данном случае готовые антитела попадают естественным путем из одного организма в другой. Пример: попадание антител матери в организм . Такой вид иммунитета может обеспечить лишь кратковременную защиту (на время существования данных антител).
Приобретенный естественный иммунитет
. Образование антител происходит в результате попадания естественным путем в организм антигенов (в результате заболевания). Формирующиеся при этом «клетки памяти» способны сохранить информацию о конкретном антигене значительное время.
Искусственный активный иммунитет
. Возникает при введении в организм искусственным путем небольшого количества антигена в виде вакцины.
Искусственный пассивный
. Возникает при введении человеку готовых антител извне. Например, при введении готовых антител против столбняка. Действие такого иммунитета непродолжительно. Особые заслуги в разработке теории иммунитета принадлежат Луи Пастеру, Эдуарду Дженнеру, И. И. Мечникову.
Иммунная система состоит из различных компонентов - органов, тканей и клеток, отнесённых к этой системе по функциональному критерию (выполнение иммунной защиты организма) и анатомофизиологическому принципу организации (органно-циркуляторный принцип). В иммунной системе выделяют: первичные органы (костный мозг и тимус), вторичные органы (селезёнка, лимфатические узлы, пейеровы бляшки и др.), а также диффузно расположенную лимфоидную ткань - отдельные лимфоидные фолликулы и их скопления. Особо выделяют лимфоидную ткань, ассоциированную со слизистыми оболочками (Mucosa-Associated Lymphoid Tussue - MALT).
Лимфоидная система - совокупность лимфоидных клеток и органов. Часто лимфоидную систему упоминают как анатомический эквивалент и синоним иммунной системы, однако это не вполне верно. Лимфоидная система является лишь частью иммунной системы: по лимфатическим сосудам клетки иммунной системы мигрируют к лимфоидным органам - месту индукции и формирования иммунного ответа. Кроме того, лимфоидную систему не следует путать с лимфатической - системой лимфатических сосудов, по которым происходит циркуляция лимфы в организме. Лимфоидная система тесно связана с кровеносной и эндокринной системами, а также с покровными тканями - слизистыми оболочками и кожей. Названные системы - основные партнёры, на которые в своей работе опирается иммунная система.
Органно-циркуляторный принцип организации иммунной системы. В организме взрослого здорового человека содержится около 10 13 лимфоцитов, т.е. примерно каждая десятая клетка тела - лимфоцит. Анатомо-физиологически иммунная система организована по органноциркуляторному принципу. Это означает, что лимфоциты не являются строго резидентными клетками, а интенсивно рециркулируют между лимфоидными органами и нелимфоидными тканями через лимфатические сосуды и кровь. Так, через каждый лимфатический узел за 1 ч проходит ≈10 9 лимфоцитов. Миграцию лимфоцитов обусловливают
специфические взаимодействия конкретных молекул на мембранах лимфоцитов и клеток эндотелия стенки сосудов [такие молекулы называют адгезинами, селектинами, интегринами, хоминг-рецепторами (от англ. home - дом, место прописки лимфоцита)]. В результате каждый орган обладает характерным набором популяций лимфоцитов и их клеток-партнёров по иммунному ответу.
Состав иммунной системы. По типу организации выделяют различные органы и ткани иммунной системы (рис. 2-1).
. Кроветворный костный мозг - место локализации стволовых кроветворных клеток (СКК).
Рис. 2-1. Компоненты иммунной системы
. Инкапсулированные органы: тимус, селезёнка, лимфатические узлы.
. Неинкапсулированная лимфоидная ткань.
-Лимфоидная ткань слизистых оболочек (MALT - MucosalAssociated Lymphoid Tissue). Независимо от локализации содержит внутриэпителиальные лимфоциты слизистой оболочки, а также специализированные образования:
◊ лимфоидная ткань, ассоциированная с пищеварительным трактом (GALT - Gut-Associated Lymphoid Tissue). В ней выделяют миндалины, аппендикс, пейеровы бляшки, lamina propria («собственная пластинка») кишечника, отдельные лимфоидные фолликулы и их группы;
◊ лимфоидная ткань, ассоциированная с бронхами и бронхиолами (BALT - Bronchus-Associated Lymphoid Tissue);
◊лимфоидная ткань, ассоциированная с женскими половыми путями (VALT - Vulvovaginal-Associated Lymphoid Tissue);
◊лимфоидная ткань, ассоциированная с носоглоткой (NALT - Nose-Associated Lymphoid Tissu e).
Особое место в иммунной системе занимает печень. В ней присутствуют субпопуляции лимфоцитов и других клеток иммунной системы, «обслуживающие» в качестве лимфоидного барьера кровь воротной вены, несущей все всасываемые в кишечнике вещества.
Лимфоидная подсистема кожи - лимфоидная ткань, ассоциированная с кожей (SALT - Skin-Associated Lymphoid Tissue) - диссеминированные внутриэпителиальные лимфоциты и региональные лимфатические узлы и сосуды лимфодренажа.
. Периферическая кровь - транспортно-коммуникационный компонент иммунной системы.
Центральные и периферические органы иммунной системы
. Центральные органы. Кроветворный костный мозг и тимус - центральные органы иммунной системы, именно в них начинается миелопоэз и лимфопоэз - дифференцировка моноцитов и лимфоцитов от СКК до зрелой клетки.
До рождения плода развитие В-лимфоцитов происходит в фетальной печени. После рождения эта функция передаётся костному мозгу.
В костном мозге проходят полные «курсы» эритропоэза (образование эритроцитов), миелопоэза (образование нейтрофилов,
моноцитов, эозинофилов, базофилов), мегакариоцитопоэза (формирование тромбоцитов), а также проходит дифференцировка ДК, NK-клеток и В-лимфоцитов. - Предшественники T-лимфоцитов для прохождения лимфопоэза мигрируют из костного мозга в тимус и слизистую оболочку пищеварительного тракта (внетимическое развитие).
. Периферические органы. В периферических лимфоидных органах (селезёнка, лимфатические узлы, неинкапсулированная лимфоидная ткань) зрелые наивные лимфоциты контактируют с антигеном и АПК. Если антигенраспознающий рецептор лимфоцита связывает комплементарный антиген в периферическом лимфоидном органе, то лимфоцит вступает на путь дальнейшей дифференцировки в режиме иммунного ответа, т.е. начинает пролиферировать и продуцировать эффекторные молекулы - цитокины, перфорин, гранзимы и др. Такую додифференцировку лимфоцитов на периферии называют иммуногенезом. В результате иммуногенеза формируются клоны эффекторных лимфоцитов, распознающих антиген и организующих деструкцию как его самого, так и периферических тканей организма, где этот антиген присутствует.
Клетки иммунной системы. В состав иммунной системы входят клетки различного происхождения - мезенхимного, экто- и энтодермального.
. Клетки мезенхимного генеза. К ним относят клетки, дифференцировавшиеся из предшественников лимфо/гематопоэза. Разновидности лимфоцитов - T, B и NK, которые в процессе иммунного ответа кооперируются с различными лейкоцитами - моноцитами/ макрофагами, нейтрофилами, эозинофилами, базофилами, а также ДК, тучными клетками и эндотелиоцитами сосудов. Даже эритроциты вносят свой вклад в реализацию иммунного ответа: транспортируют иммунные комплексы «антиген-антитело-комплемент» в печень и селёзенку для фагоцитоза и разрушения.
. Эпителий. В состав некоторых лимфоидных органов (тимус, некоторые неинкапсулированные лимфоидные ткани) входят эпителиальные клетки эктодермального и энтодермального происхождения.
Гуморальные факторы. Помимо клеток, «иммунная материя» представлена растворимыми молекулами - гуморальными факторами. Это продукты B-лимфоцитов - антитела (они же иммуноглобулины) и растворимые медиаторы межклеточных взаимодействий - цитокины.
ТИМУС
В тимусе (thymus) проходит лимфопоэз значительной части T-лимфоцитов («Т» происходит от слова «Thymus»). Тимус состоит из 2 долей, каждая из которых окружена капсулой из соединительной ткани. Перегородки, идущие от капсулы, разделяют тимус на дольки. В каждой дольке тимуса (рис. 2-2) выделяют 2 зоны: по периферии - корковая (cortex), в центре - мозговая (medulla). Объём органа заполнен эпителиальным каркасом (эпителий), в котором располагаются тимоциты (незрелые Т-лимфоциты тимуса), ДК и макрофаги. ДК расположены преимущественно в зоне, переходной между корковой и мозговой. Макрофаги присутствуют во всех зонах.
. Эпителиальные клетки своими отростками обхватывают лимфоциты тимуса (тимоциты), поэтому их называют «nurse cells» (клетки-«сиделки» или клетки-«няньки»). Эти клетки не только поддерживают развивающиеся тимоциты, но также продуцируют

Рис. 2-2. Строение дольки тимуса
цитокины ИЛ-1, ИЛ-3, ИЛ-6, ИЛ-7, LIF, GM-CSF и экспрессируют молекулы адгезии LFA-3 и ICAM-1, комплементарные молекулам адгезии на поверхности тимоцитов (CD2 и LFA-1). В мозговой зоне долек расположены плотные образования из скрученных эпителиальных клеток - тельца Гассаля (тельца тимуса) - места компактного скопления дегенерирующих эпителиальных клеток.
. Тимоциты дифференцируются из костномозговых СКК. Из тимоцитов в процессе дифференцировки образуются Т-лимфоциты, способные распознавать антигены в комплексе с MHC. Однако большинство Т-лимфоцитов либо не сможет обладать этим свойством, либо будет распознавать аутоантигены. Для предотвращения выхода таких клеток на периферию в тимусе инициируется их элиминация путем индукции апоптоза. Таким образом, в норме в циркуляцию из тимуса выходят только клетки, способные распознавать антигены в комплексе со «своими» MHC, но при этом не индуцирующие развитие аутоиммунных реакций.
. Гематотимический барьер. Тимус сильно васкуляризован. Стенки капилляров и венул образуют гематотимический барьер на входе в тимус и, возможно, на выходе из него. Зрелые лимфоциты выходят из тимуса либо свободно, так как каждая долька имеет эфферентный лимфатический сосуд, выносящий лимфу в лимфатические узлы средостения, либо путём экстравазации через стенку посткапиллярных венул с высоким эндотелием в корково-мозговой области и/или через стенку обычных кровеносных капилляров.
. Возрастные изменения. К моменту рождения тимус полностью сформирован. Он густо заселён тимоцитами в течение всего детства и до момента полового созревания. После пубертата тимус начинает уменьшаться в размерах. Тимэктомия у взрослых не приводит к серьёзным нарушениям иммунитета, поскольку в детстве и подростковом возрасте создаётся необходимый и достаточный пул периферических T-лимфоцитов на всю оставшуюся жизнь.
ЛИМФАТИЧЕСКИЕ УЗЛЫ
Лимфатические узлы (рис. 2-3) - множественные, симметрично расположенные, инкапсулированные периферические лимфоидные органы бобовидной формы, размером от 0,5 до 1,5 см в длину (при отсутствии воспаления). Лимфатические узлы через афферентные (приносящие) лимфатические сосуды (их несколько на каждый узел) дренируют тка-

Рис. 2-3. Строение лимфатического узла мыши: а - корковая и мозговая части. В корковой части расположены лимфатические фолликулы, от которых в мозговую часть отходят мозговые тяжи; б - распределение T- и B-лимфоцитов. Тимусзависимая зона выделена розовым цветом, тимуснезависимая зона - жёлтым. T-лимфоциты поступают в паренхиму узла из посткапиллярных венул и вступают в контакт с фолликулярными дендритными клетками и B-лимфоцитами
невую жидкость. Таким образом, лимфатические узлы - «таможня» для всех веществ, в том числе для антигенов. Из анатомических ворот узла вместе с артерией и веной выходит единственный эфферентный (выносящий) сосуд. В итоге лимфа попадает в грудной лимфатический проток. Паренхима лимфатического узла состоит из T-клеточной, B-клеточной зон и мозговых тяжей.
. B-клеточная зона. Корковое вещество разделено соединительнотканными трабекулами на радиальные секторы и содержит лимфоидные фолликулы, это B-лимфоцитарная зона. Строма фолликулов содержит фолликулярные дендритные клетки (ФДК), формирующие особое микроокружение, в котором происходит уникальный для B-лимфоцитов процесс соматического гипермутагенеза вариабельных сегментов генов иммуноглобулинов и отбор наиболее аффинных вариантов антител («созревание аффинности антител»). Лимфоидные фолликулы проходят 3 стадии развития. Первичный фолликул - мелкий фолликул, содержащий наивные B-лимфоциты. После того как B-лимфоциты вступают в иммуногенез, в лимфоидном фолликуле появляется герминативный (зародышевый) центр, содержащий интенсивно пролиферирующие B-клетки (это происходит примерно через 4-5 дней после активной иммунизации). Это вторичный фолликул. По завершении иммуногенеза лимфоидный фолликул существенно уменьшается в размере.
. T-клеточная зона. В паракортикальной (T-зависимой) зоне лимфатического узла расположены T-лимфоциты и интердигитальные ДК (они отличаются от ФДК) костномозгового происхождения, которые презентируют антигены T-лимфоцитам. Через стенку посткапиллярных венул с высоким эндотелием происходит миграция лимфоцитов из крови в лимфатический узел.
. Мозговые тяжи. Под паракортикальной зоной расположены содержащие макрофаги мозговые тяжи. При активном иммунном ответе в этих тяжах можно видеть множество зрелых B-лимфоцитов - плазматические клетки. Тяжи впадают в синус мозгового вещества, из которого выходит эфферентный лимфатический сосуд.
СЕЛЕЗЁНКА
Селезёнка - относительно большой непарный орган массой около 150 г. Лимфоидная ткань селезёнки - белая пульпа. Селезёнка - лимфоцитарная «таможня» для антигенов, попавших в кровь. Лимфоциты

Рис. 2-4. Селезёнка человека. Тимусзависимая и тимуснезависимая зоны селезёнки. Скопление T-лимфоцитов (зелёные клетки) вокруг артерий, вышедших из трабекул, образует тимусзависимую зону. Лимфатический фолликул и окружающая его лимфоидная ткань белой пульпы формируют тимуснезависимую зону. Так же как и в фолликулах лимфатических узлов, здесь присутствуют B-лимфоциты (жёлтые клетки) и фолликулярные дендритные клетки. Вторичный фолликул содержит герминативный центр с быстроделящимися В-лимфоцитами, окружёнными кольцом малых покоящихся лимфоцитов (мантией)
селезёнки накапливаются вокруг артериол в виде так называемых периартериолярных муфт (рис. 2-4).
T-зависимая зона муфты непосредственно окружает артериолу. B-клеточные фолликулы расположены ближе к краю муфты. Артериолы селезёнки впадают в синусоиды (это уже красная пульпа). Синусоиды заканчиваются венулами, собирающимися в селезёночную вену, несущую кровь в воротную вену печени. Красную и белую пульпу разделяет диффузная маргинальная зона, населенная особой популяцией В-лимфоцитов (В-клетки маргинальной зоны) и особыми макрофагами. Клетки маргинальной зоны являются важным связующим звеном между врождённым и приобретённым иммунитетом. Здесь происходит самый первый контакт организованной лимфоидной ткани с возможными патогенами, циркулирующими в крови.
ПЕЧЕНЬ
Печень выполняет важные иммунные функции, что вытекает из следующих фактов:
Печень - мощный орган лимфопоэза в эмбриональном периоде;
Аллогенные трансплантаты печени отторгаются менее интенсивно, чем другие органы;
Толерантность к вводимым перорально антигенам можно индуцировать только при нормальном физиологическом кровоснабжении печени и не удаётся индуцировать после операции по созданию портокавальных анастомозов;
Печень синтезирует белки острой фазы (СРБ, MBL и др.), а также белки системы комплемента;
В печени содержатся разные субпопуляции лимфоцитов, в том числе уникальные лимфоциты, сочетающие признаки T- и NK-клеток (NKT-клетки).
Клеточный состав печени
Гепатоциты формируют паренхиму печени и содержат очень мало молекул MHC-I. Молекулы MHC-II гепатоциты в норме почти не несут, однако их экспрессия может возрастать при заболеваниях печени.
Клетки Купфера - макрофаги печени. Они составляют около 15% от общего числа клеток печени и 80% всех макрофагов организма. Плотность макрофагов выше в перипортальных областях.
Эндотелий синусоидов печени не имеет базальной мембраны - тонкой внеклеточной структуры, состоящей из разных типов коллагенов и других белков. Эндотелиальные клетки формируют монослой с просветами, через которые лимфоциты могут непосредственно контактировать с гепатоцитами. Кроме того, эндотелиальные клетки экпрессируют различные рецепторы-«мусорщики» (scavenger-рецепторы).
Лимфоидная система печени, кроме лимфоцитов, содержит анатомический отдел циркуляции лимфы - пространства Диссе. Эти пространства с одной стороны непосредственно контактируют с кровью синусоидов печени, а с другой - с гепатоцитами. Лимфоток в печени значителен - не менее 15-20% всего лимфотока организма.
Звёздчатые клетки (клетки Ито) расположены в пространствах Диссе. Они содержат жировые вакуоли с витамином А, а также характерные для гладкомышечных клеток α-актин и десмин. Звёздчатые клетки могут трансформироваться в миофибробласты.
ЛИМФОИДНАЯ ТКАНЬ СЛИЗИСТЫХ ОБОЛОЧЕК И КОЖИ
Неинкапсулированная лимфоидная ткань слизистых оболочек представлена глоточным лимфоидным кольцом Пирогова-Вальдейера, пейеровыми бляшками тонкой кишки, лимфоидными фолликулами аппендикса, лимфоидной тканью слизистых оболочек желудка, кишечника, бронхов и бронхиол, органов мочеполовой системы и других слизистых оболочек.
Пейеровы бляшки (рис. 2-5) - групповые лимфатические фолликулы, расположенные в lamina propria тонкой кишки. Фолликулы, точнее T-клетки фолликулов, примыкают к кишечному эпителию под так называемыми M-клетками («М» от Membranous, эти клетки не имеют микроворсинок), являющимися «входными воротами» пейеровой бляшки. Основная масса лимфоцитов расположена в B-клеточных фолликулах с зародышевыми центрами. T-клеточные зоны окружают фолликул ближе к эпителию. B-лимфоциты составляют 50-70%, T-лимфоциты - 10-30% всех клеток пейеровой бляшки. Основная функция пейеровых бляшек - поддержание иммуногенеза B-лимфоцитов и их дифференци-

Рис. 2-5. Пейерова бляшка в стенке кишки: а - общий вид; б - упрощённая схема; 1 - энтероциты (эпителий кишки); 2 - М-клетки; 3 - T-клеточная зона; 4 - B-клеточная зона; 5 - фолликул. Масштаб между структурами не выдержан
ровка в плазматические клетки, продуцирующие антитела - преимущественно секреторные IgA. Продукция IgA в слизистой оболочке кишки составляет более 70% общей ежедневной продукции иммуноглобулинов в организме - у взрослого человека около 3 г IgA каждый день. Более 90% всего синтезируемого организмом IgA экскретируется через слизистую оболочку в просвет кишки.
Внутриэпителиальные лимфоциты. Помимо организованной лимфоидной ткани в слизистых оболочках есть и одиночные внутриэпителиальные T-лимфоциты, диссеминированные среди эпителиальных клеток. На их поверхности экспрессирована особая молекула, обеспечивающая адгезию этих лимфоцитов к энтероцитам, - интегрин α Е (CD103). Порядка 10-50% внутриэпителиальных лимфоцитов составляют TCRγδ + CD8αα + T-лимфоциты.
Иммунная система различает «свое» и «чужое» и уничтожает потенциально опасные чужеродные молекулы и клетки из организма. Иммунная система также обладает способностью выявлять и разрушать патологически измененные клетки собственных тканей. Любая молекула, распознаваемая иммунной системой, рассматривается как антиген (АГ).
Кожа, роговица и слизистая оболочка дыхательных путей, желудочно-кишечного тракта образуют физический барьер, являющийся первой линией защиты человеческого тела. Некоторые из этих барьеров обладают активными иммунными функциями:
- Внешний, ороговевший эпидермис: кератиноциты кожи секретируют антимикробные пептиды (дефензины), а сальные и лотовые железы секретируют субстанции, подавляющие микробов. В коже присутствуют многие другие иммунные клетки.
- Слизистая оболочка дыхательных путей, желудочно-кишечного и мочеполового трактов: слизистая оболочка содержит антимикробные вещества, такие как лизоцим, лактоферины и секреторный иммуноглобулин A (SlgA).
При нарушении иммунных барьеров реализуются 2 типа иммунитета: врожденный и приобретенный. Многие молекулярные составляющие участвуют и во врожденном и в приобретенном иммунитете.
Врожденный иммунитет
Врожденный (естественный) иммунитет не требует предварительной встречи с антигенами. Таким образом, он немедленно отвечает на агрессора. Он распознает главным образом молекулы широко представленных антигенов, а не специфичных данному организму или клетке. Его компонентами являются:
- фагоцитирующие клетки,
- антигенпредставляющие клетки,
- естественные клетки-киллеры,
- полиморфно-ядерные лейкоциты.
Фагоцитирующие клетки (нейтрофилы и моноциты крови, макрофаги и дендритические клетки тканей) поглощают и разрушают вторгающиеся АГ Атака фагоцитирующих клеток облегчается, если антигены прикрываются антителами (AT), что является частью приобретенного иммунитета или когда белки комплемента (являющиеся частью менее специфичной врожденной системы защиты) опсонизируют АГ. АГ-презентирующие клетки представляют фрагменты поглощенных АГ Т-лимфоцитам и являются частью приобретенного иммунитета. Естественные клетки-киллеры уничтожают клетки, зараженные вирусом и клетки некоторых опухолей.
Приобретенный иммунитет
Приобретенный иммунитет требует предварительной встречи с антигенами, т.е. ему требуется время для развития после первичной встречи с новым агрессором. Далее следует быстрый ответ. Система запоминает предшествующие контакты и является АГ-специфичной. Ее компонентами являются:
- Т-клетки.
- В клетки.
Приобретенный иммунитет, полученный от определенных Т-клеточных иммунных реакций, называется клеточно-опосредованным иммунитетом. Иммунитет, полученный от В-клеточных реакций, называют гуморальным иммунитетом, т.к. В клетки секретируют растворимые Ag-специфичные антитела. В-клетки и Т-клетки работают вместе, уничтожая инородные элементы. Некоторые из этих клеток не уничтожают инородные элементы напрямую, а вместо этого активируют другие лейкоциты, которые распознают и уничтожают инородные вещества.
Иммунный ответ
Успешная иммунная защита требует активации, регуляции и реализации иммунного ответа.
Активация . Иммунная система активируется чужеродным АГ, который распознается циркулирующими AT или клеточными рецепторами. Эти рецепторы могут быть высокоспецифичными или низкоспецифичными. Низкоспецифичные рецепторы распознают общие группы лигандов, входящих в структуру факторов патогенности микроорганизмов, такие как липополисахариды грамотрицательных бактерий, пептидогликаны грамположительных бактерий, бактериальные флагеллины, неметилированные цитозин-гуанозин динуклеотиды (CpG-мотивы) и вирусные двухполосные ДНК. Активация также возникает, если комплексы АТ-АГ и комплемент-микроорганизм связываются с поверхностными клеточными рецепторами для Fc-фрагмента IgG или для фрагментов комплемента С.
Распознанный АГ, комплексы АГ-АТ или комплемент-микроорганизм подвергаются фагоцитозу. Большинство микроорганизмов уничтожаются в результате фагоцитоза, другие (например, микобактерия) подавляют способность фагоцитов полностью уничтожать их, хотя и не препятствуют поглощению. В таких случаях цитокины, продуцируемые Т-лимфоцитами, в частности IgG, у (ИФН-γ), стимулируют выработку фагоцитами литических ферментов и других микробицидных веществ, которые убивают микроорганизмы.
Пока АГ подвергается быстрому фагоцитозу и полностью разрушается (не частый случай), работает приобретенный иммунный ответ. Он берет начало в селезенке для АГ, циркулирующих в кровотоке, в лимфоузлах для тканевых АГ и в лимфоидных тканях, ассоциированных с слизистой оболочкой (например, миндалевидные железы, аденоиды, Пейеровы бляшки) для АГ слизистой оболочки. Например, дендритные клетки Лангерганса подвергают фагоцитозу антигены в коже и мигрируют в местные лимфоузлы, где производные от АГ пептиды экспрессированы на клеточной поверхности молекул класса II главного комплекса гистосовместимости (МНС), которые представляют пептид CD4 хелперным клеткам (ТН). Когда Т-хелперная клетка работает с комплексом МНС-пептид и получает костимуляторные сигналы, она активируется и экспрессирует рецепторы для ИЛ-2 цитокина и секретирует некоторые цитокины. Каждый набор Т-хелперных клеток секретирует различные комбинации веществ, таким образом, влияя на природу иммунного ответа.
Регуляция . Иммунный ответ должен регулироваться для предотвращения чрезвычайного вреда организму (например, анафилаксия, значительные повреждения тканей). Регуляторные Т-клетки помогают контролю иммунного ответа посредством секреции цитокинов иммуносупрессоров, таких как ИЛ-10 и трансформирующий фактор роста-β (ТФР-β) или же посредством малоизученного механизма клеточного контакта. Данные регуляторные клетки предотвращают возникновение аутоиммунного ответа, и по-видимому, способствуют реализации ответов на несобственные (чужеродные) АГ.
Завершение . Завершение иммунного ответа происходит тогда, когда АГ отделен или удален из организма. Без антигенной стимуляции прекращается секреция цитокинов и активированные цитотоксические клетки подвергаются апоптозу. Апоптоз маркирует клетку для незамедлительно следующего фагоцитоза, препятствующего потере клеточного содержимого и развитию воспаления. Т-клетки и В-клетки, дифференцированные в клетки памяти, избегают данной участи.
Гериатрическая составляющая
С возрастом иммунная система становится менее эффективной, а именно:
- Ослабевает ее способность распознавать собственное от чужеродного, увеличивая частоту аутоиммунных нарушений.
- Макрофаги уничтожают бактерии, раковые клетки и другие АГ менее интенсивно, что объясняет увеличение случаев онкологий среди пожилых людей.
- T-клетки не способны быстро реагировать на АГ.
- Уменьшается количество лимфоцитов, способных реагировать на новые АГ.
- Стареющее тело вырабатывает меньшее количество комплемента в ответ на бактериальные инфекции.
- Меньшее количество AT вырабатывается в ответ на АГ, a AT обладают меньшей способностью прикрепляться к АГ, что объясняет повышение частоты случаев пневмоний, гриппа, инфекционного эндокардита и столбняка и повышенного риска смертельного исхода данных патологий у пожилых. Эти изменения могут также отчасти объяснять неэффективность вакцинаций среди пожилых людей.
Компоненты иммунной системы
Иммунная система состоит из клеточных и молекулярных компонентов, которые работают совместно с целью уничтожения АГ.
Антигенпрезентирующие клетки
Хотя некоторые антигены могут непосредственно стимулировать иммунный ответ,Т-зависимый приобретенный иммунный ответ обычно требует присутствия АГ-презентирующих клеток (АПК), которые представляют пептиды АГ в комплексе с молекулами МНС. Внутриклеточные АГ (например, вирусные) могут быть преобразованы и представлены рецепторам CD8 Тс-лимфоцитов любыми ядерными клетками. С помощью кодирующих их белков, которые вмешиваются в данный процесс, некоторые вирусы (например, цитомегаловирус) избегают уничтожения. Внутриклеточный АГ должен быть преобразован в пептид и представлен в комплексе с молекулами МНС II класса на поверхности АПК для распознавания клетками хелперами, несущими CD4 клеткам.
Моноциты в кровотоке являются предшественниками тканевых макрофагов. Моноциты мигрируют в ткани, где через 8 ч они развиваются в макрофаги под воздействием колониестимулирующего фактора макрофагов (М-КСФ), секретируемого различными клеточными типами (например, клетками эндотелия, фибробластными клетками).
Макрофаги активируются IFN-y и колониестимулирующим фактором гранулоцитов-макрофагов (ГМ-КСФ). Активированные макрофаги уничтожают внутриклеточные организмы и секретируют ИЛ-1 и фактор некроза опухолей-альфа (TNF-α). Эти цитокины потенцируют секрецию IFN-γ и GM-CSF и увеличивают экспрессию молекул адгезии на поверхности эндотелиоцитов, облегчая приток лейкоцитов к очагу инфекции и разрушение патогенного фактора. В зависимости от профиля экспрессии гена макрофаги были классифицированы в субтипы.
Дендритные клетки присутствуют в коже (клетки Лангерганса), лимфоузлах, в тканях всего организма. Дендритные клетки в коже - пограничные АПК, они захватывают АГ, доставляют его в локальные лимфоузлы, где активируют Т-лимфоциты.
Однако у них есть рецепторы к Fc-фрагменту IgG и комплементу, что позволяет им связывать иммунные комплексы и представлять их В-лимфоцитам зародышевых центров вторичных лимфоидных органов.
Полиморфно-ядерные лейкоциты
Полиморфно-ядерные лейкоциты (ПЯЛ) также называются гранулоцитами, т.к. их цитоплазма содержит специфические гранулы.
Они присутствуют в циркулирующей крови и имеют сегментированное ядро, за исключением тучных клеток, которые постоянно присутствуют в тканях и функционально подобны циркулирующим базофилам.
Нейтрофилы составляют 40-70% всех лейкоцитов; они являются первой линией защиты в борьбе с инфекцией. У зрелых нейтрофилов полупериод существования составляет от 2 до 3 дней. Во время острого воспалительного процесса (например, инфекционного) нейтрофилы, реагируя на хемотаксические факторы, покидают кровяное русло и выходят в ткани. Их цель - фагоцитировать и разрушить патогенные факторы. Микроорганизмы уничтожаются в том случае, когда фагоциты продуцируют литические ферменты и активные формы кислорода (супероксид, гипохлористая кислота) или запускают высвобождение содержимого гранул (дефенсины, протеазы, повышающие проницаемость тканей бактерицидные протеины, лактоферрин и лизоцим). Также высвобождаются ДНК и гистоны, и они вместе с содержимым гранул, таким как эластаза, образуют волокна в окружающих тканях, что может способствовать уничтожению бактерий и локализации ферментной активности.
Базофилы составляют менее 5% лейкоцитов и сходны с тучными клетками, хотя принадлежат к разным клеточным линиям. И те и другие клетки имеют высокоаффинные рецепторы для IgE. Когда эти клетки сталкиваются с определенным АГ, этот АГ перекрестно связывает соседние бивалентные молекулы IgE, что вызывает дегрануляцию клеток с высвобождением готовых медиаторов воспаления и образованием новых медиаторов (лейкотриены, простагландины, тромбоксаны).
Тучные клетки имеются в разных тканях организма. У тучных клеток слизистых оболочек гранулы содержат триптазу и хондроитин сульфат, а если клетка локализована в соединительной ткани, то в ее гранулах содержатся триптаза, химаза, гепарин. При выбросе этих медиаторов формируется защитный острый воспалительный ответ. Дегрануляция может быть запущена анафилатоксином, фрагментами комплемента C3a и C5a.
Цитотоксичные лейкоциты
К цитотоксическим лейкоцитам относятся:
- Естественные клетки-киллеры.
- Лимфокин-активированные киллеры.
Естественные клетки-киллеры (ЕКК) . Типичные ЕКК составляют от 5 до 15% моно-нуклеарных клеток периферической крови. Они имеют круглое ядро и гранулированную цитоплазму. ЕКК индуцируют апоптоз в инфицированных и анормальных клетках различными путями. Как клетки врожденного ответа у них отсутствуют антигенспецифичные рецепторы и иммунологическая память.
Типичные ЕКК очень важны при контроле за мутирующими клетками, т.к. они экспрессируют и активирующие и ингибиторные рецепторы. Активирующие рецепторы ЕКК распознают множество лигандов клеток-мишеней (например, относящуюся к МНС класса I цепь A и цепь В. Ингибиторные рецепторы ЕКК распознают молекулы МНС класса I. ЕКК уничтожают свои мишени только при отсутствии сильного сигнала ингибиторных рецепторов. Наличие молекул МНС класса I (в норме экспрессированных на ядросодержащих клетках) на клетках предотвращает их разрушение; а отсутствие указывает на то, что клетка инфицирована каким-то вирусом, ингибирующим экспрессию МНС или что потеряла экспрессию МНС, потому что рак изменил данную клетку. Больные с дефицитом ЕКК особенно чувствительны к герпесной инфекции и человеческому папилломавирусу (вирусу папилломы человека).
ЕКК также секретируют некоторые цитокины; они являются основным источником IFN-γ. Секретируя IFN-γ, ЕКК могут влиять на приобретенную иммунную систему, способствуя дифференциации (дифференцировке) клеток хелперов 1-го типа (Тн1) и ингибируя хелперы 2-го типа (Тн2).
Лимфокин-активированные киллеры (LAK) . Некоторые лимфоциты развиваются в очень мощные лимфокин-активированные киллеры (LAK), способные уничтожать широкий спектр опухолевых клеток и аномальных лимфоцитов (например, инфицированные определенными вирусами). Эти клетки не только составляют уникальный клеточный подтип лимфоцитов, они - феноминальны. Предшественники LAK гетерогенны, но изначально могут быть классифицированы как ЕКК-подобные (чаще всего) или Т-лимфоцитоподобные клетки.
Лимфоциты
2 важнейшими типами лимфоцитов являются:
- В-лимфоциты, которые созревают в костном мозге.
- Т-лимфоциты, которые созревают в тимусе.
Они не различаются с точки зрения морфологии, но имеют разные иммунные функции. Они отличаются друг от друга АГ-специфическими поверхностными рецепторами, молекулами, называемыми кластерами дифференцировки (CD), которые присутствуют или отсутствуют в определенном клеточном подтипе. Идентифицировано более 300 CD. Каждый лимфоцит распознает специфический АГ посредством поверхностных рецепторов.
В-лимфоциты . От 5 до 15% лимфоцитов крови - это В-лимфоциты. Они также присутствуют в селезенке, лимфоузлах на слизистой оболочке лимфоидных тканей. В-клетки могут представлять АГ Т-клеткам, но их первоочередной функцией является развитие в клетки плазмы, производящие и секретирующие антитела (AT). Больные с В-клеточными иммунодефицитами (например, Х-связанная агаммаглобулинемия) особенно восприимчивы к рецидивирующим бактериальным инфекциям.
После случайной реаранжировки генов, кодирующих lg, В-лимфоциты способны узнавать практически бесконечное число уникальных АГ. Реаранжировка генов последовательно происходит во время развития В-лимфоцитов в красном костном мозге. Процесс начинается с коммитированной стволовой клетки, проходит стадии про-В- и пре-В-лимфоцитов и завершается незрелым В-лимфоцитом. Если этот незрелый В-лимфоцит взаимодействует с АГ, то может произойти инактивация (развитие толерантности) или элиминация (апоптоз) этой клетки. Незрелые В-лимфоциты, которые не подверглись инактивации или элиминации, могут продолжить развитие до стадии зрелого молодого В-лимфоцита, покинуть красный костный мозг и переместиться в периферические лимфоидные органы, где может произойти их встреча с АГ. Их ответ на АГ происходит в 2 этапа:
- Первичный иммунный ответ. Когда зрелые молодые В-лимфоциты впервые сталкиваются с АГ, эти клетки претерпевают бласттрансформацию, клональную пролиферацию и дифференцировку в клетки памяти, которые отреагируют на такой же АГ в будущем, или в зрелые АТ-продуцирующие плазмоциты. До продукции AT существует латентный период в несколько дней. Потом продуцируются только IgM. Вначале продуцируются только IgM. После взаимодействия с Т-лимфоцитами в В-лимфоцитах может иметь место дальнейшая реаранжировка генов lg, что переключает синтез на IgG, IgA или IgE.
- Вторичный иммунный ответ (анамнестический, усиленный). Когда В-клетки памяти и Т-хелперные клетки повторно встречаются с тем же АГ. В-клетки памяти быстро пролиферируют, дифференцируются в зрелые плазмоциты, быстро синтезируют и выделяют большое количество AT (главным образом IgG, т.к. Т-лимфоциты индуцируют переключение синтеза именно этого изотипа) в кровь и другие ткани, где AT могут реагировать с АГ. Таким образом, после повторной встречи с АГ иммунный ответ более быстрый и эффективный. Т-лимфоциты.
Существует 3 основных типа Т-лимфоцитов:
- Хелперные.
- Регуляторные.
- Цитотоксические.
Более зрелые Т-лимфоциты экспрессируют CD4 или CD8, а также АГ-связывающие lg-подобные рецепторы, называемые Т-клеточными рецепторами (TCR). Гены, кодирующие TCR, подобно иммуноглобулиновым генам, реаранжируются. В результате достигается определенная специфичность и аффинность при контакте с молекулами МНС, представленными на мембране АПК и связанными с пептидами АГ. Число специфичных связей у Т-лимфоцитов практически бесконечно.
Для активации Т-лимфоцита TCR связывается или с комплексом АГ-МНС или со вспомогательными молекулами; в противном случае Т-лимфоцит останется неактивированным или погибнет в результате апоптоза. Некоторые вспомогательные молекулы ингибируют ранее активированные Т-лимфоциты и таким образом завершают иммунный ответ. Полиморфизм гена CTLA-4 ассоциирован с некоторыми аутоиммунными патологиями.
Т-Хелперными (Тн) клетками обычно являются CD4, но могут быть и CD8. Они дифференцируются из Тн0-клеток в одни из следующих:
Каждый тип клеток секретирует определенные цитокины. Имеются различные общие схемы продукции цитокинов, которые и определяют для Тн-клеточные функциональные фенотипы. Тн2-клетки способны до определенного уровня регулировать в сторону понижения функциональную активность друг друга, что приводит к доминированию Тн1- или Тн2-ответа.
Различие между разными типами β-клетками клинически достоверно. Например, Тн1-ответ преобладает при туберкулоидной лепре, а Тн2-ответ преобладает при лепроматозной лепре. Тн1-ответ характерен для некоторых аутоиммунных патологий, а Тн2-ответ способствует продукции IgG и развитию аллергических заболеваний, а также помогает В-клеткам секретировать антитела при некоторых аутоиммунных патологиях (например, болезнь Грейвса, миастения гравис). Для больных с иммунодефицитными состояниями характерны дефективные Тн 17-клетки (например, синдром гипер IgE ),такие больные наиболее восприимчивы к инфекциям, вызванным Candida albicans и Staphylococcus aureus.
Регуляторные Т-клетки. Опосредуют супрессию иммунных ответов и обычно экспрессируют фактор транскрипции Fox3. В данном процессе участвуют профессиональные клеточные подсемейства CD4 CD8, они либо секретируют цитокины, обладающие иммуносупрессивными свойствами, либо подавляют иммунный ответ, механизм подавления пока плохо изучен и требует прямого контакта между клетками. У больных с функциональными мутациями в Foxp3 развиваются аутоиммунная патология, синдром IPEX (иммунодисрегуляция, полиэндокринопатия, энтеропатия, Х-связанная).
Цитотоксическими Т(Тс)-клетками, как правило, являются CD8, но могут быть и CD4; они необходимы для уничтожения внутриклеточных патогенов, в частности, вирусов.
Тс-клетки проходят 3 стадии развития:
- Клетка-предшественник, которая при соответствующей стимуляции дифференцирует в Тс-клетку.
- Дифференцированная клетка-эффектор, способная уничтожать мишени.
- Клетка памяти, находящаяся в покое (уже не стимулированная), но готовая выполнять эффекторную функцию после повторной стимуляции оригинальным сочетанием АГ-МНС.
Полностью активированные Тс клетки, подобно ЕКК, способны уничтожать инфицированную клетку-мишень, индуцируя апоптоз.
Тс клетки могут быть:
- Изогенными: произведенными в ответ на собственные (аутологичные) клетки, модифицированные вирусной инфекцией или другими чужеродными белками.
- Аллогенными: произведенными в ответ на клетки, экспрессирующие чужеродные продукты МНС (например, при органных трансплантациях когда молекулы донорского МНС отличаются от МНС реципиента) Некоторые Тс-клетки могут направленно распознавать чужеродные МНС (прямой путь); другие могут распознавать фрагменты чужеродного МНС, представленные собственными молекулами МНС реципиента трансплантата (непрямой путь).
Антитела
AT функционируют как антигенный рецептор на поверхности В-клеток и в ответ на АГ секретируются плазмоцитами. AT распознают специфические конфигурации на поверхности АГ (например, белки, полисахариды, нуклиновые кислоты). AT и АГ точно подходят друг другу, поскольку их формы и другие поверхностные свойства (например, нагрузка) комплементарны. Одна и та же молекула AT может перекрестно реагировать с соответствующим АГ, если их эптопы достаточно схожи с эпитопами исходного АГ.
Структура . AT состоят из 4 полипептидных цепей (2 идентичных тяжелых цепей и 2 идентичных легких цепей), связанных дисульфидными связями для производства Y конфигурации. И тяжелая и легкая цепи имеют варабильный (V) и константный (С) участки.
V - Вариабельные участки располагаются на аминотерминальных концах верхней части Y; они называются вариабельными, потому что содержат различные аминокислоты, которые и определяют специфичность lg. Гипервариабельные регионы содержат идиотипические детерминанты, с которыми связываются определенные естественные (антиидиотипические) AT; эта связь может помочь в регуляции В-гуморального ответа. В-лимфоциты могут изменять изотип тяжелой цепи продуцируемого lg, но сохраняет тяжелые цепи V-региона и полностью легкую цепь, в связи с чем сохраняется АГ-специфичность.
С-регион состоит из относительно постоянной последовательности аминокислот, которая характерна для каждого lg-изотипа.
Аминотерминальный (вариабильный) конец AT связывается с АГ и формирует комплекс АГАТ. АГ-связывающая часть lg (Fab) состоит из легкой цепи и фрагмента тяжелой и включает V-регион молекулы lg (смешанная часть).
Классы антител . Антитела подразделяются на 5 классов:
Данные классы различаются по типу тяжелой цепи; существует также 2 типа легких цепей (к и А). Все 5 классов lg имеют либо к- либо λ-легкие цепи.
IgM является первым AT, которое формируется после встречи с новым АГ. Он состоит из 5 Y-молекул (10 тяжелых и 10 легких цепей), связанных простой связью. IgM циркулирует преимущественно во внутрисосудистом пространстве; он связывается и агглютинирует АГ и может активировать комплемент, что облегчает фагоцитоз. IgM представляют собой изо гемагглютинины и многие AT к грамотрицательным микроорганизмам. Мономер IgM является АГ-рецептором на поверхности В-лимфоцитов. У больных с синдромом гипер-lgM имеется дефект генов, участвующих во включении конкретного класса антител (например, генов, кодирующих CD40 или CD154); поэтому уровни IgA, IgM и IgE у них низкие или вовсе отсутствуют, а уровни циркулирующих IgM зачастую высоки.
IgG преобладающий изотип lg; он циркулирует и во внутри- и во внесосудистом пространствах. IgG - первично циркулирующий lg, появляющийся после реиммунизации (при вторичном иммунном ответе) и являющийся доминантным изотипом в коммерческой у глобулиновой продукции. IgG защищает организм от бактерий, вирусов,токсинов, и это единственный изотип lg, который проходит через плацентарный барьер. Вот почему данный класс антител важен как протектор новорожденных, но пато генные IgG антитела, в случае присутствия в организме будущей матери могут спровоцировать серьезное патологическое состояние плода.
Выделяют 4 субкласса IgG: IgG1, LgG2, lgG3, lgG4, пронумерованные по убыванию концентрации IgG в сыворотке. Субклассы IgG отличаются главным образом по их способности активировать комплемент; IgG1 и LgG3 наиболее эффективны, lgG2 менее эффективен и LgG4 неэффективен. IgG1 и IgG3 являются эффективными медиаторами клеточной цитотоксичности, опосредованной антителами; lgG4 и lgG2 менее эффективны в этом плане.
IgA присутствует на поверхностях слизистых оболочек, в сыворотке и в секретах (слюна, слезная жидкость, секрет респираторного, желудочно-кишечного и мочеполового трактов, молозиво), где обеспечивает начальную антибактериальную и антивирусную защиту. J-цепь связывает IgA в димер - формируется молекула секреторного IgA. Секреторный IgA синтезируется плазмацитами в субэпителиальной части слизистой оболочки ЖКТ и дыхательных путей. Избирательный дефицит IgA относительно распространен, но не имеет большого значения с точки зрения клиницистов, поскольку существует перекрестная функциональность между другими классами антител.
IgD коэкспрессирован с IgM на поверхности молодых В-лимфоцитов. Имеют ли эти 2 класса различные функции, и если да, то насколько -пока неясно. Они могут быть просто примером молекулярной деградации. Уровни IgD в сыворотке очень низки, а функция циркулирующего IgD неизвестна.
Реактанты острой фазы
Реактанты острой фазы - белки плазмы, уровень которых резко повышается или в некоторых случаях понижается при инфекционных процессах или поражении тканей. Наиболее значительно повышается С-реактивный белок и маннозосвязывающий лектин (который фиксирует белки комплемента и играет роль опсонина), α 1 -транспортный белок кислого гликопротеина и сывороточный компонент амилоида СРВ и СОЭ часто измеряются; повышение уровней является неспецифическим признаком инфекции или воспаления. Повышенный фибриноген является главной причиной повышения ESR.
Многие реактанты острой фазы вырабатываются в печени. В совокупности они помогают ограничивать повреждение тканей, повышают резистентность организма к инфекциям, способствуют репарации тканей и прекращению воспаления.
Цитокины
Цитокины - это полипептиды, секретируемые иммунными и другими клетками после их взаимодействия со специфическим АГ, эндотоксином и другими цитокинами. Основные группы цитокинов включают интерфероны:
- интерфероны;
- факторы некроза опухолей (TNF-α, лимфотоксины-α,лимфотоксины-β);
- интерлейкины (IL);
- хемокины;
- трансформирующие факторы роста;
- гемопоэтические колониестимулирующие факторы (CSF).
Хотя лимфоцит начинает секрецию цитокина после взаимодействия со специфическим АГ, сами по себе цитокины не являются АГ-специфическими.
Цитокины передают сигналы через рецепторы клеточной поверхности. Например, рецептор И/1-2 состоит из 3 цепей: α, β и γ. Аффинность рецептора для ИЛ-2 будет высокой, если экс-прессированы все 3 цепи, средней, если экс-прессированы только β- и γ-цепи, и низкой, если экспрессирована только α-цепь. Мутации либо делеции уцепи составляют основу Х-цепленного тяжелого комбинированного иммунодефицита.
Хемокины индуцируют хемотаксис и миграцию лейкоцитов. Выделяют 4 субсемейства, отличающихся количеством разделяющих аминокислот между первыми двумя цистеиновыми остатками. Хемокиновые рецепторы (CCR5 на Т-лимфоцитах памяти, моноцитах/макрофагах, дендритических клетках; CXCR4 на остальных Т-лимфоцитах) являются корецепторами для проникновения ВИЧ (вируса иммунодефицита человека) в клетку.
Лейкоцитарные антигены человека
Система лейкоцитарных антигенов человека (HLA) локализуется в 6-й хромосоме. Эта хромосома кодирует молекулы клеточной поверхности.
Молекулы МНС класса I присутствуют на поверхности всех ядерных клеток в качестве трансмембранных гликопротеинов; после того как данные молекулы денатурируются и расщепляются, их поглощают тромбоциты. Нормальная молекула класса I состоит из тяжелой цепи а, связанной с р2-молекулой микроглобулина. Тяжелая цепь состоит из двух связанных пептидных доменов, lg-подобного домена, трансмембранного участка и цитоплазматического конца. Тяжелая цепь молекулы МНС I класса кодируется генами HLA-A, -В или С-локусов. Лимфоциты, реагирующие на молекулы МНС I класса, экс-прессируют молекулы CD8 и выполняют эффекторные функции, заключающиеся в способности распознавать любые инфицированные клетки. Поскольку любая ядросодержащая клетка экс-прессирует молекулы МНС класса I, все инфицированные клетки являются антигенпрезентирующими для CD8 положительных Т лифоцитов (CD8 связывается с неполиморфным участком тяжелой цепи класса I). Некоторые гены МНС класса I кодируют неклассические МНС-молекулы, например HLA-G и HLA-E (которые представляют пептиды некоторым рецепторам ЕКК).
Молекулы МНС класса II обычно представлены только на профессиональных АГ-презентирующих клетках, эпителиальных клетках тимуса и активированных (но не покоящихся) Т-клетках; большинство ядросодержащих клеток могут быть стимулированы для экспрессии молекул МНС класса II интерфероном (IFN)-γ. Молекулы МНС класса I состоят из двух полипетидных (а и (3) цепей; у каждого пептида имеется пептид-связывающий участок, lg-подобный участок и трансмембранный регион с цитоплазматическим хвостиком. Обе полипептидные цепи кодируются генами регионов HLA-DP, -DQ, or -DR 6-й хромосомы. Лифоциты, реагирующие на молекулы МНС класса II, экспрессируют CD4 и часто являются Т-клетками-хелперами.
Регион МНС класса III кодирует некоторые молекулы, значимые при воспалении.
Отдельные определенные с помощью серологического типирования антигены, кодируемые генами локусов класса I и II, имеют стандартные обозначения. Аллели, определенные с помощью ДНК-секвенирования, содержат в обозначении название гена, далее идет звездочка, потом цифры, обозначающие аллельную группу (часто соответствующую серологически выявленному антигену, кодируемому данным аллелем), потом - двоеточие и цифры, обозначающие данный аллель. Иногда в обозначении аллеля имеются после двоеточия дополнительные цифры для обозначения аллельных вариантов, кодирующих идентичные белки, и после второго двоеточия добавлены цифры для обозначения полиморфизмов в интронах или же в 5" или 3" нетранслируемых регионах.
Молекулы МНС класса I и II являются самыми иммуногенными антигенами и распознаются в процессе отторжения аллогенного трансплантата. Сильнейшей детерминантой является HLA-DR, затем идут HLA-B и -А.Три данные локуса поэтому и являются важнейшими при подборе подходящего (тканесовместимого) донора реципиенту.
Система комплемента
Система комплемента - каскад ферментов, облегчающих борьбу с инфекционным процессом. Эта система связывает врожденный и приобретенный иммунитет путем:
- Увеличения ответа антител (AT) и иммунологической памяти.
- Лизинга чужеродных молекул.
- Удаления иммунных комплексов. Компоненты системы комплемента выполняют множество биологических функций.
Активация белков комплемента: существует 3 пути активации комплемента:
- классический,
- лектиновый (маннозосвязывающий лектин-MBL),
- альтернативный.
Компоненты классического пути обозначаются буквой С и цифрой, обозначающей очередность их идентификации. Компоненты альтернативного пути часто обозначаются буквенно (например, фактор В, фактор D) или отдельным названием (например, пропердин).
Классический путь. Активация классического пути - АТ-зависимый процесс, который начинается после взаимодействия С1 с комплексом АГ-lgM или АГ-lgG, или АТ-независимый процесс, когда полианионы (гепарин, протамин, ДНК или РНК апоптотических клеток), грамотрицательные бактерии или связанный С-реактивный белок реагируют непосредственно с С1. Этот путь регулируется С1-ингибитором (C1-INM). Наследственная ангиоэдема ассоциирована с генетической недостаточностью C1-INH.
Лектиновый путь (маннозосвязывающий лектин) - АТ-независимый процесс; он начинается, когда MBL-сывороточный белок связывается с маннозой, фруктозой.
Альтернативный путь начинается с прилипания компонентов микробной клеточной поверхности или lg к небольшому количеству С3. Этот путь регулируется пропердином, фактором Н, фактором, ускоряющим некроз.
Эти 3 пути в конце концов сходятся в одну точку, когда С3-конвертаза превращает С3 в С3a и С3b. Расщепление С3 может привести к образованию комплекса атаки на мембрану (MAC), цитотоксическому компоненту системы комлпемента. MAC является причиной лизиса чужеродных клеток.
Больные с дефицитом компонентов комплемента часто подвержены рецидивирующим бактериальным инфекциям, в частности при отсутствии компонента С3. Дефекты С1 и С4 ассоциированы с системной красной волчанкой.
Биологическая активность . Компоненты системы комплемента выполняют и другие биологические функции, которые реализуются рецепторами комплемента (CR) на различных типах клеток.
- CR1 (CD35) способствует фагоцитозу и участвует в выведении иммунных комплексов.
- CR2 (CD21) регулирует продукцию AT В-лимфоцитами и является рецептором вируса Эпштейна - Барр.
- CR3 (CDllb/CD18), SR4(CDllc/CD18) и рецепторы Clq играют роль в фагоцитозе.
- С3а, С5а и С4а (слабо) проявляют анафилатическую активность. Они вызывают дегрануляцию тучных клеток, ведущую к повышению проницаемости сосудов и сокращению гладкой мускулатуры.
- С3b работает в качестве опсонина покрывая микроорганизмы и тем самым усиливая их фагоцитоз.
- C3d усиливает продукцию AT В-лимфоцитами.
- С5а является хемоэттрактантом нейтрофилов.Он контролирует активность нейтрофилов и моноцитов и может стать причиной повышенного слипания клеток, дегрануляции и высвобождения внутриклеточных ферментов из гранулоцитов, продукцию токсических метаболитов кислорода и другие действия, связанные с клеточным мета болизмом.
Функции иммунной системы
Основной функцией иммунной системы является надзор за макромолекулярным и клеточным постоянством организма, защита организма от всего чужеродного. Иммунная система вместе с нервной и эндокринной системами регулируют и контролируют все физиологические реакции организма, тем самым, обеспечивая жизнедеятельность и жизнеспособность организма. Иммунокомпетентные клетки являются обязательным элементом воспалительной реакции и во многом определяют характер и ход её течения. Важной функцией иммунокомпетентных клеток является контроль и регуляция процессов регенерации тканей.
Свою основную функцию иммунная система осуществляет через развитие специфических (иммунных) реакций, в основе которых лежит способность распознавания "своего" и "чужого" и последующая элиминация чужеродного. Появляющиеся в результате иммунной реакции специфические антитела составляют основу гуморального иммунитета, а сенсибилизированные лимфоциты являются основными носителями клеточного иммунитета.
Иммунная система обладает феноменом "иммунологической памяти", который характеризуется тем, что повторный контакт с антигеном вызывает ускоренное и усиленное развитие иммунного ответа, что обеспечивает более эффективную защиту организма по сравнению с первичной иммунной реакцией. Эта особенность вторичной иммунной реакции лежит в основе смысла вакцинации, которая успешно защищает от большинства инфекций. Следует отметить, что иммунные реакции не всегда выполняют только защитную роль, они могут быть причиной иммунопатологических процессов в организме и обусловливать целый ряд соматических заболеваний человека.
Структура иммунной системы
Иммунная система человека представлена комплексом лимфомиелоидных органов и лимфоидной ткани, ассоциированной с дыхательной, пищеварительной и мочеполовой системами. К органам иммунной системы относятся: костный мозг, тимус, селезёнка, лимфатические узлы. В состав иммунной системы, помимо перечисленных органов, также входят миндалины носоглотки, лимфоидные (пейеровы) бляшки кишечника, многочисленные лимфоидные узелки, расположенные в слизистых оболочках желудочно-кишечного тракта, дыхательной трубки, урогенитальных путей, диффузная лимфоидная ткань, а также лимфоидные клетки кожи и межэпителиальные лимфоциты.
Главным элементом иммунной системы являются лимфоидные клетки. Общее число лимфоцитов у человека составляет 1012 клеток. Вторым важным элементом иммунной системы являются макрофаги. Кроме этих клеток, в защитных реакциях организма участвуют гранулоциты. Лимфоидные клетки и макрофаги объединены понятием иммунокомпетентные клетки.
В иммунной системе выделяют Т-звено и В-звено или Т-систему иммунитета и В-систему иммунитета. Основными клетками Т-системы иммунитета являются Т-лимфоциты, основными клетками В-системы иммунитета В-лимфоциты. К главным структурным образованиям Т-системы иммунитета относятся тимус, Т-зоны селезёнки и лимфатических узлов; В-системы иммунитета – костный мозг, В-зоны селезёнки (центры размножения) и лимфатических узлов (кортикальная зона). Т-звено иммунной системы ответственно за реакции клеточного типа, В-звено иммунной системы реализует реакции гуморального типа. Т-система контролирует и регулирует работу В-системы. В свою очередь, В-система способна оказывать влияние на работу Т-системы.
Среди органов иммунной системы различают центральные органы и периферические органы. К центральным органам относятся костный мозг и тимус, к периферическим – селезёнка и лимфатические узлы. В костном мозге из стволовой лимфоидной клетки происходит развитие В-лимфоцитов, в тимусе из стволовой лимфоидной клетки происходит развитие Т-лимфоцитов. По мере созревания Т- и В-лимфоциты покидают костный мозг и тимус и заселяют периферические лимфоидные органы, расселяясь соответственно в Т- и В-зонах.
Из чего состоит кровь?
Кровь состоит из форменных элементов (или клеток крови) и плазмы. На плазму приходится 55-60% всего объема крови, клетки крови составляют соответственно 40-45%.
Плазма
Плазма представляет собой слегка желтоватую полупрозрачную жидкость с удельным весом 1,020-1,028 (удельный вес крови 1,054-1,066) и состоит из воды, органических соединений и неорганических солей. 90-92% составляет вода, 7-8% - белки, 0,1% - глюкоза и 0,9% - соли.
Клетки крови
Эритроциты
В плазме крови взвешены красные кровяные тельца, или эритроциты. Эритроциты многих млекопитающих и человека представляют двояковогнутые диски, не имеющие ядер. Диаметр эритроцитов человека равен 7-8 µ, а толщина - 2-2,5 µ. Образование эритроцитов происходит в красном костном мозге, в процессе созревания они теряют ядра, а затем поступают в кровь. Средняя продолжительность жизни одного эритроцита составляет примерно 127 дней, затем эритроцит разрушается (преимущественно в селезенке).
Гемоглобин
Молекулы гемоглобина из старых эритроцитов в селезенке и печени подвергаются расщеплению, атомы железа используются снова, а гем разрушается и выделяется печенью в виде билирубина и других желчных пигментов. Ядерные эритроциты могут появиться в крови после больших кровопотерь, а также при нарушении нормальных функций ткани красного костного мозга. У взрослого мужчины в 1 мм3 крови содержится около 5 400 000 эритроцитов, а у взрослой женщины - 4 500 000 - 5 000 000. У новорожденных детей эритроцитов больше - от 6 до 7 млн в 1 мм3. Каждый эритроцит содержит около 265 млн молекул гемоглобина - красного пигмента, переносящего кислород и углекислоту. Подсчитано, что ежесекундно образуется около 2,5 млн эритроцитов и столько же разрушается. А так как в каждом эритроците содержится 265·106 молекул гемоглобина, то ежесекундно образуется примерно 650·1012 молекул такого же гемоглобина.
Гемоглобин состоит из двух частей: белковой - глобина и железосодержащей - гема. В капиллярах легких кислород диффундирует из плазмы в эритроциты и соединяется с гемоглобином (Hb), образуя оксигемоглобин (HbO2): Hb+O2 « HbO2. В капиллярах тканей в условиях низкого парциального давления кислорода комплекс HbO2 распадается. Гемоглобин, соединенный с кислородом, называется оксигемоглобином, а гемоглобин, отдавший кислород - восстановленным гемоглобином. Некоторое количество СO2 переносится кровью в форме непрочного соединения с гемоглобином - карбооксигемоглобина.
Лейкоциты
Кровь содержит пять видов белых кровяных телец, или лейкоцитов, - бесцветных клеток, содержащих ядро и цитоплазму. Они образуются в красном костном мозгу, лимфатических узлах и селезенке. Лейкоциты лишены гемоглобина и способны к активному амебоидному движению. Лейкоцитов меньше, чем эритроцитов - в среднем около 7 000 на 1 мм3, но число их колеблется в пределах от 5 000 до 9 000 (или 10 000) у разных людей и даже у одного и того же человека в разное время суток: меньше всего их рано утром, а больше всего - после полудня. Лейкоциты делятся на три группы: 1) зернистые лейкоциты, или гранулоциты (их цитоплазма содержит гранулы), среди них различают нейтрофилы, эозинофилы и базофилы; 2) незернистые лейкоциты, или агранулоциты, - лимфоциты; 3) моноциты.
Тромбоциты
Есть еще одна группа форменных элементов - это тромбоциты, или кровяные пластинки, - наименьшие из всех клеток крови. Они образуются в костном мозгу. Количество их в 1 мм3 крови колеблется от 300 000 до 400 000. Они играют важную роль в начале процесса свертывания крови. У большинства позвоночных тромбоциты представляют собой небольшие овальные клетки, имеющие ядро, тогда как у млекопитающих - это мельчайшие дисковидные пластинки. При кровотечениях их них выделяется вещество серотонин, вызывающее сужение сосудов. Содержание тромбоцитов увеличивается при мышечной работе (миогенный тромбоцитоз). В тромбоцитах обнаружены железо и медь, а также дыхательные ферменты.
Не пропустите - всё интересное рубрики "ЗДОРОВЬЕ " --> !










